Griffith Transformation
In 1928, Frederick Griffith performed the first experiment to prove that DNA was the hereditary material. Griffith selected the bacterium that causes pneumonia, Diplococcus pneumoniae. Two strains of the bacterium namely virulent strain and avirulent strain were used for the study.
Virulence required the presence of a polysaccharide capsule around the bacterium. The avirulent mutants lacked this capsule. The colonies of avirulent bacteria did not have capsule and appeared rough. They were designated R. In contrast, the virulent form produced colonies that appeared smooth, so it was designated S. Virulent forms were of different types and each had a characteristic polysaccharide capsule (called I S, II S, III S, etc.), which is genetically inherited and is immunologically distinct from other forms.
Virulent bacterium of a particular capsule type (say II S) can mutate to a non-encapsulated, nonvirulent form (II R, because it derives from a Type II cell) and vice versa. This happens at a very low frequency (in less than one in a million cells), but it is inherited when it does occur. However, the II R cell line cannot mutate to a III S virulent form (Figure 1.13 (a)).
Figure 1.13 (a) Diplococcus pneumoniae–virulent and avirulent strains (b) Griffith experiment
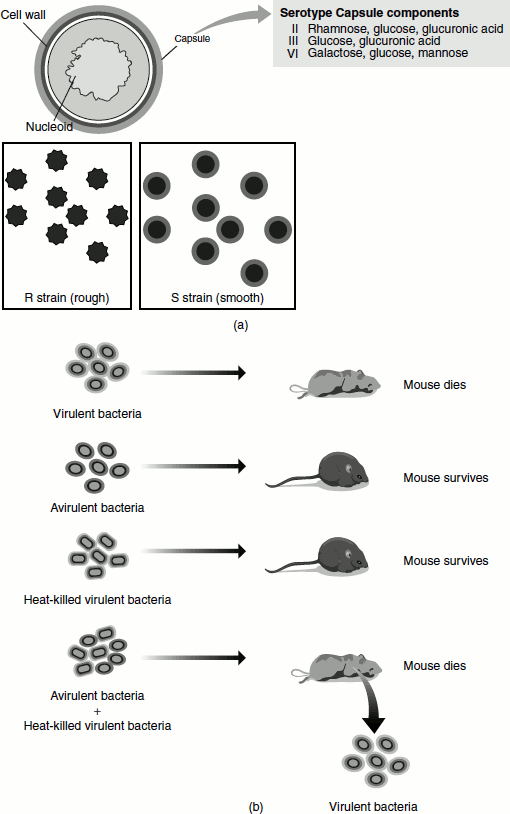
When attenuated encapsulated Type III S cells were injected into mice, the mice did not develop pneumonia. Similarly, when II R cells were injected into mice, the mice did not develop the illness. When live Type III S cells were injected, it resulted in the death of the animal.
Griffith mixed Pneumococcus Type II R with attenuated II S cells. Both the strains mentioned when injected alone did not produce the disease. Therefore, no disease was expected from the mixed injections, as neither strain was virulent. However, many of the mice given mixed injections developed pneumonia and died. When the blood of the animal was analysed, they all contained living virulent Type III S cells. These cells could not have arisen from the Type II R cells by mutations (they would have produced Type II S cells), and the Type III S cells were attenuated. This proves that some factors must have passed from the dead III S cells to the live II R ones, which would have enabled them to make a capsule of the Type III transforming them to Type III S. Griffith called the factor as ‘transforming principle’ and the process as genetic transformation (Figure 1.13 (b)).
Oswald Avery, Colin MacLeod and Maclyn McCarty Experiment
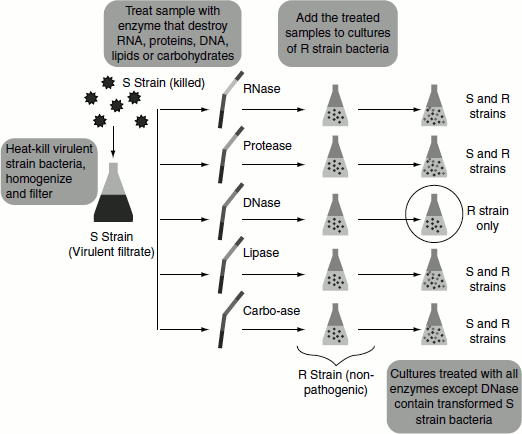
The nature of the Griffith’s transforming principle was studied by Oswald Avery, together with his colleagues Colin MacLeod and Maclyn McCarty, of Columbia University, New York. These scientists followed Griffith’s experiment but with some changes. In order to identify the transforming principle, they isolated the R-II transformed S-III cells from the dead mice. The S-III cells were then lysed and the cellular contents of the cell were subjected to different enzyme treatments. To five tubes containing the cellular lysate, one of the following enzymes was added: RNase, an enzyme that destroys RNA; protease, an enzyme that destroys protein; DNase, an enzyme that destroys DNA; lipase, an enzyme that destroys Lipids; or a combination of enzymes that break down carbohydrates. To the sixth tube, no enzyme was added and used as the control tube.
If the ‘transforming agent’ was, for example, protein—the transforming agent would be destroyed in the test tube containing protease, but not the others. Thus, whatever the transforming agents was, the liquid in one of the tubes would no longer be able to transform the S. pneumoniae strains. The scientists observed that the liquid from the tubes that received RNase, protease, lipase and the carbohydrate-digesting enzymes was still able to transform the R strain of pneumonia into the S strain. However, the liquid that was treated with DNase completely lost the ability to transform the bacteria. Thus, it was apparent that the ‘transforming agent’ in the liquid was DNA (Figure 1.14).
To further prove their finding, the scientists took a liquid extracted from attenuated S. pneumoniae (S strain) and subjected it to extensive preparation and purification, isolating only the pure DNA from the mixture. This pure DNA was also able to transform the R strain into the S strain and generate pathogenic S. pneumoniae. These results provided powerful evidence that DNA, and not protein, was actually the genetic material inside of living cells.
Disadvantages of the Experiment
Avery’s experiments had several complicating factors.
- It was not clear in the minds of all microbiologists that transformation really was a genetic phenomenon.
- There were doubts about the specificity of the deoxyribonuclease enzyme that he used to inactivate the transforming principle.
- It was believed that the added enzyme contained trace amounts of a contaminating protease and hence was also able to degrade protein.
Figure 1.14 Oswald Avery, Colin MacLeod and Maclyn McCarty experiment
These uncertainties necessitated the need for a second experiment to provide more information on the chemical nature of the genetic material.
Hershey and Chase Experiment
In 1952, two scientists named Alfred Hershey and Martha Chase performed an entirely different type of genetic experiment using bacteriophages to prove that DNA is the carrier of genetic information. Bacteriophages (or just phage, an extremely small virus) are viruses that infect bacterial cells. They use the host cell machinery and transform it into a factory for producing more phages. Scientist of the time knew that the phage itself does not enter the bacterium during an infection. Rather, a small amount of material is injected into the bacteria and this material must contain all of the information necessary to build more phages. Thus, this injected substance is the genetic material of the phage.
Hershey and Chase devised a very simple experiment using a technique called radioactive labelling to determine which molecule, whether DNA, RNA or protein, acted as the genetic material in phages.
Figure 1.15 Hershey and chase experiment
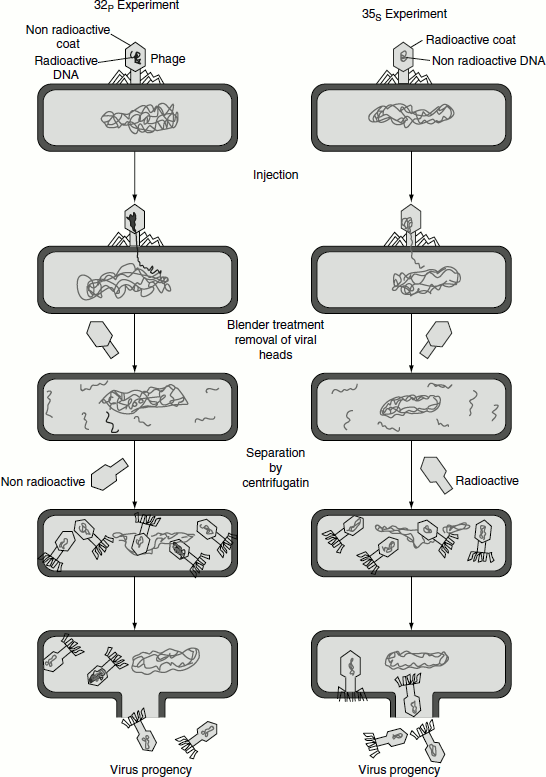
The chemical make-up of protein and of DNA is quite different. DNA contains phosphorus but proteins do not; on the other hand, proteins usually contain sulphur but DNA does not. By specifically labelling the phosphorus and sulphur atoms with radioisotopes, Hershey and Chase were able to distinguish between the protein and the DNA of the phage and determine whether either or both were injected into the bacterial cell during the course of infection.
Two batches of isotopically labelled bacteriophage particles were prepared. One was labelled with 32P in the phosphate groups of the DNA and the other with 35S in the sulphur-containing groups of the amino acids of the protein coat called capsid, of the virus. The two batches of labelled phages were allowed to infect the bacterial cells. After a short time interval, the suspension of phage-infected bacterial cells was agitated in a blender that sheared the viral capsids from the bacteria. The blended mixture was then centrifuged. This separated the bacterial cells from the empty viral ghost consisted only capsid. The cells infected with the 32P-labelled phage were found to contain 32P, indicating that the labelled viral DNA has entered the cell, while there was no radioactivity detected in the viral ghost (Figure 1.15).
The opposite occurred when 35S-labelled phage infected a bacterial culture. The cells infected with 35S-labelled phage were found to have no radioactivity after blender treatment but their viral ghosts remained suspended in the supernatant after centrifugation contained 35S. This proved that 35S-labelled phage protein did not enter the bacterial cell. This experiment proved that only the DNA from the phage entered the bacterial cell and dictated the information required for the production of progeny phages; in other words, the DNA is the carrier of genetic information (Figure 1.15).
A small amount of protein did enter the bacterial cell in the course of infection. However, this was not involved in the production of new bacteriophage. This fact was demonstrated by repeating the experiment with bacteria stripped of their cell walls (protoplasts). If protoplasts were infected with 32P phage DNA free of protein, virulent phages were produced. If the purified 32P was first treated with DNAase, no progeny phage was produced. This clearly confirmed that the labelled DNA contained all the information necessary to produce new virus particles.